1.在NIID模型中,GGC重复突变可诱导含polyG的核内包涵体形成
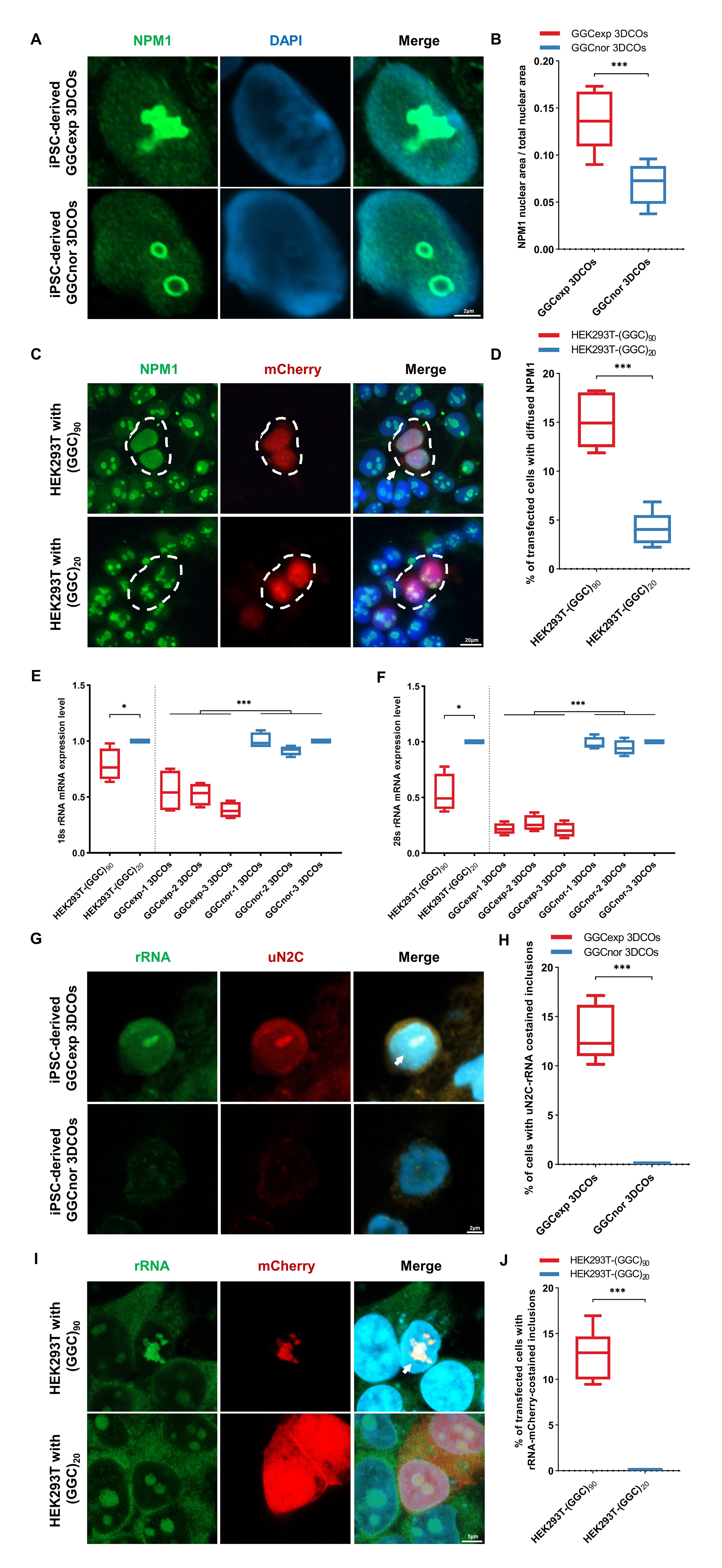
为了研究NOTCH2NLC基因5’非编码区GGC重复扩增对人脑的影响,作者对来自3例具有NOTCH2NLC GGC重复扩增的NIID患者(GGCexp)和3个正常对照(GGCnor)的皮肤成纤维细胞进行重编程,并将其分化为3D脑类器官(3DCOs)及细胞模型。免疫荧光结果发现,相较于GGCnor,GGCexp组中有更多SQSTM1泛素阳性核内包体沉积。针对NOTCH2NLC基因上游开放序列(uN2C)的免疫荧光显示,SQSTM1泛素阳性核内包体出现更多的uN2CpolyG共定位。且SQSTM1-21阳性核内包体中,uN2C polyG共定位最多,表明NOTCH2NLC GGC重复序列可以产生含有polyG的蛋白,这些蛋白出现在SQSTM1和泛素阳性的核内包体中。免疫荧光结果还发现重复RNA也存在于NIID的核内包涵体中。作者对90-DIV 3DCOs和30-DIV神经元中的NOTCH2NLC进行了定量,免疫印迹结果显示,NOTCH2NLC在GGCexp和GGCnor 3DCOs中的表达无显著差异。这些结果表明毒性RNA和翻译产物功能获得可能参与了NIID的发病过程。
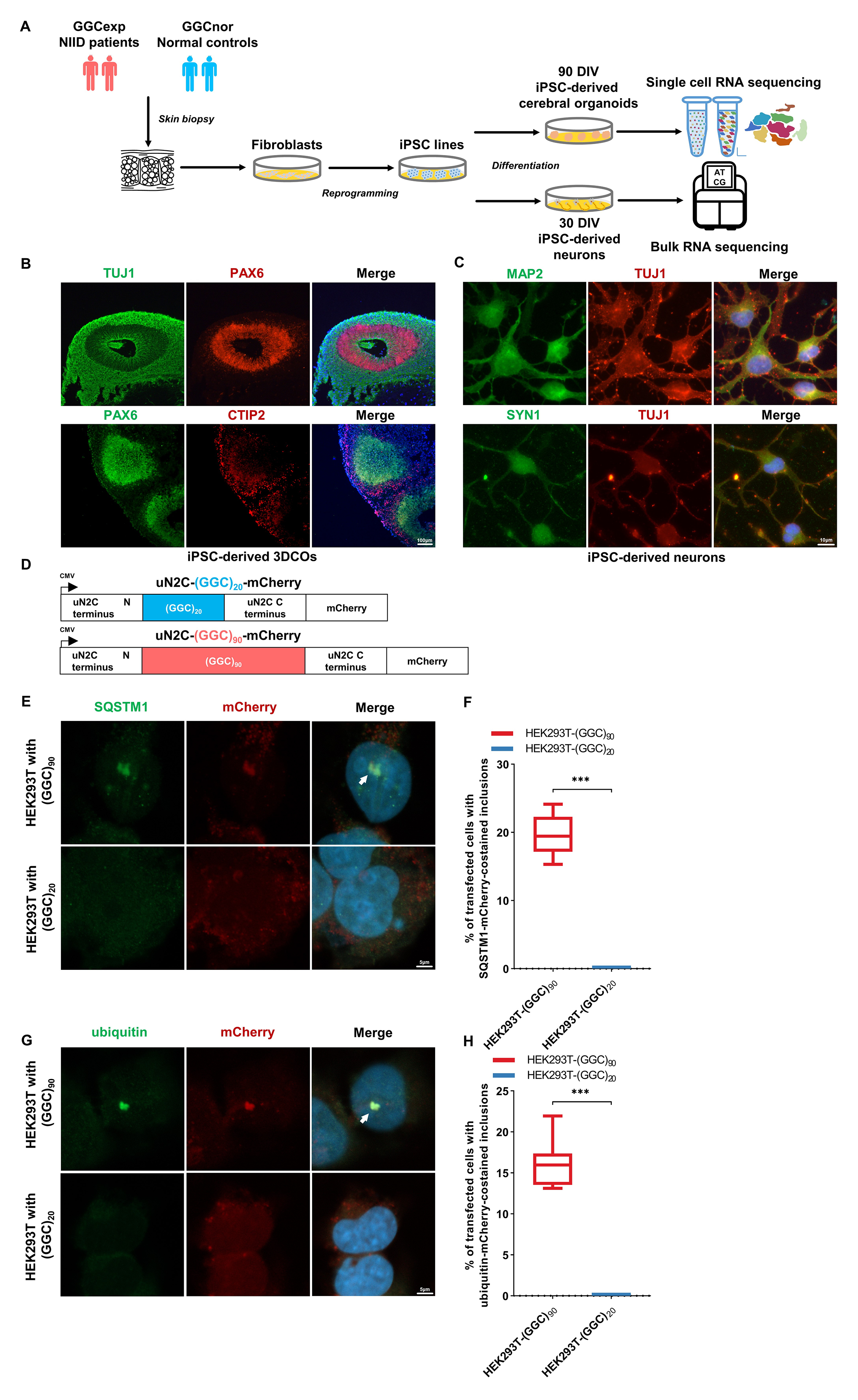
图1 患者特异性ipsc衍生的3D脑类器官、2D神经元和NIID细胞模型的生成和病理特征
图2 NOTCH2NLC基因GGC重复扩增可在脑类器官和细胞模型中诱导polyG阳性核内包体的形成
2.NOTCH2NLC GGC重复可增加自噬通量,激活整合应激反应(ISR)
SQSTM1是选择性自噬的靶点,被发现广泛地存在于NIID的核内包涵体中。免疫印迹结果显示,GGCexp 3DCOs中SQSTM1表达水平显著降低。自噬标志物LC3B在GGCexp 3DCOs和神经元中的表达显著增加。作者使用溶酶追踪器检测活细胞的溶酶体状态。在GGCexp神经元中发现溶酶-示踪剂标记的酸性成分体积和数量增加。腺病毒的免疫荧光显示GGCexp神经元中自噬小体和自噬溶酶体的数量和体积显著增加,这些结果表明NOTCH2NLC GGC重复可以增强自噬通量。同时作者定量了3DCOs和神经元中ISR标记物的表达水平,发现在GGCexp 表达增加,表明在GGCexp模型中ISR被激活。这些结果表明NOTCH2NLC GGC重复增强了自噬通量,激活了EIF2α-ISR-ATF4通路,从而促进了NOTCH2NLC GGC重复的发病机制。
图3 NOTCH2NLC GGC重复序列可引起自噬通量增加,激活综合应激反应
3.Bulk转录组测序和单细胞测序显示GGC重复序列可能影响核糖体生成及翻译
对NIID患者(GGCexp-1和GGCexp-2)和正常对照(GGCnor-1和GGCnor-2)的30-DIV神经元中进行了RNA测序,GGCexp中的下调基因主要富集在翻译和核糖体结构组成途径中。WGCNA分析发现,富集在翻译、核糖体和核糖体亚单位途径中的模块在GGCexp神经元中表达水平也显著较低。
对NIID患者和正常对照的3DCOs进行了单细胞转录组测序(scRNA-seq)。鉴定出八种细胞类型:成熟神经元(MNs)、未成熟神经元(iMNs);放射状胶质细胞(RGCs);神经祖细胞(NPCs);成神经细胞(NBs);少突胶质前体细胞(OPCs);髓鞘化雪旺氏细胞(MSCs);谷氨酸能神经元(GNs)。
GGCexp 3DCOs显示未成熟神经元(iMNs)比例较低,RGCs比例较高。伪时间轨迹分析结果发现GGCexp 3DCO在发育iMNs时分支更少,轨迹更短,而在发育RGCs时轨迹更复杂,提示NOTCH2NLC GGC重复序列可能影响3DCOs的生长。功能富集分析显示,GGCexp 3DCOs中下调基因主要参与核糖体、细胞质翻译、核糖体结构组成和细胞质核糖体。GGCexp中MNs、iMNs、RGCs、NPCs和NBs中的下调基因在细胞质翻译、核糖体和核糖体结构组成通路中一致富集。GSVA分析发现在GGCexp 3DCOs的主要细胞类型中“核糖体”和“细胞质翻译”基因的富集评分较低,表明在这些细胞中存在翻译和核糖体功能障碍。
图4 Bulk测序和单细胞测序显示,NOTCH2NLC基因中的GGC重复扩增可能影响ipsc衍生模型中的核糖体生物发生
4.GGC重复诱导核仁应激,损害核糖体的生物发生,抑制全细胞翻译
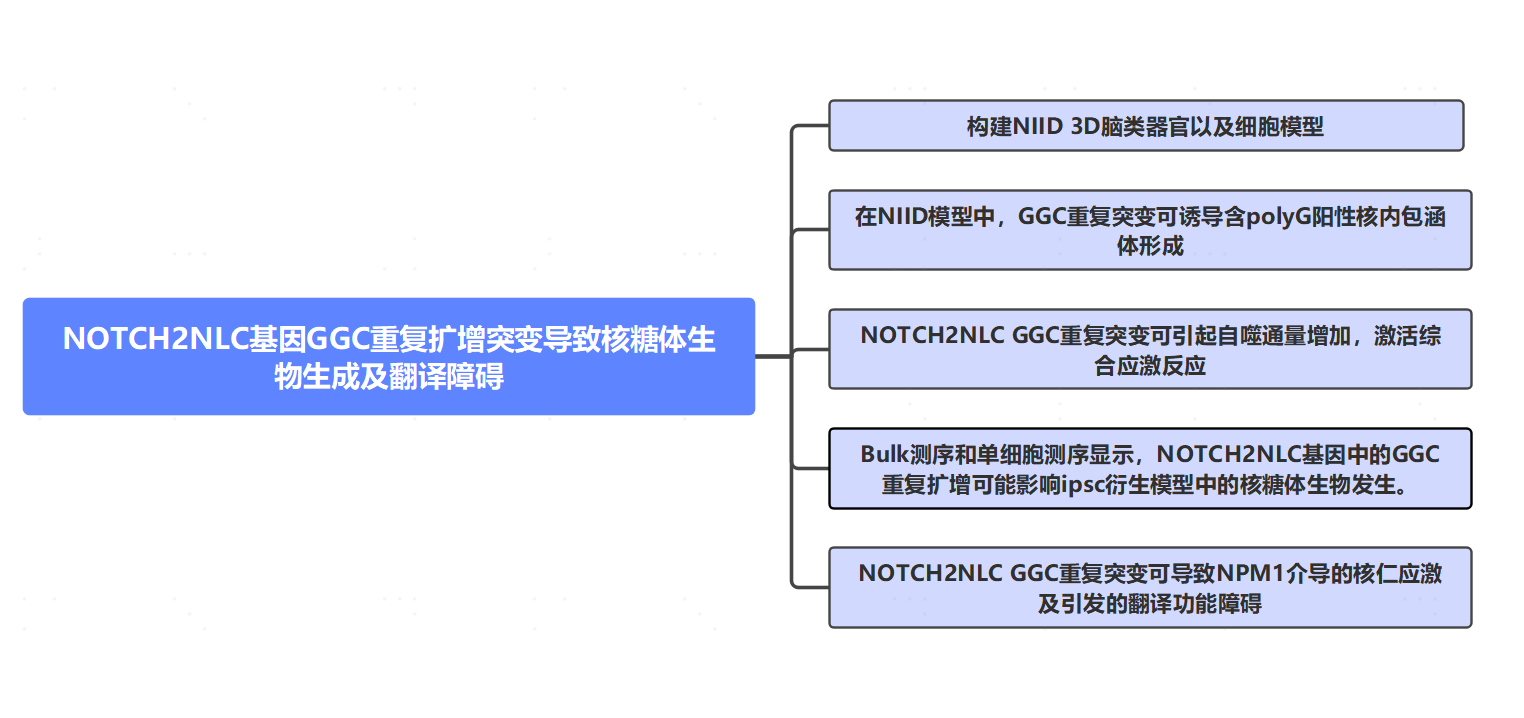
在真核细胞中,核仁的主要功能是快速合成大小核糖体亚基,参与核糖体RNA的转录、加工以及核糖体的组装。当核糖体的生成被破坏时,细胞进入核仁应激的状态,而NOTCH2NLC GGC重复序列可能导致编码核糖体蛋白的mRNA表达减少。免疫荧光显示,转染uN2C-23 (GGC)90-mCherry质粒的HEK293T细胞,显示出更明显的NPM1核质易位,提示NOTCH2NLC GGC重复序列可在NIID细胞模型中引起核仁应激和NPM1易位。
rRNA是核糖体生物发生的另一个组成部分。作者通过RT-qPCR检测18S和28S rRNA的表达水平发现其表达水平明显降低。此外,在转染uN2C-(GGC)90-10 mCherry质粒的HEK293T细胞中也显示18S和28S rRNA表达水平显著降低。这些结果表明NOTCH2NLC GGC重复序列可以影响NIID细胞模型中rRNA的表达。转染uN2C-(GGC)90-mCherry质粒的HEK293T细胞的免疫荧光显示更多的mCherry-rRNA包含体,表明核内包体可以在NIID细胞模型中隔离rRNA。
图5 NOTCH2NLC GGC重复序列在NIID细胞模型中可诱导核仁应激和核糖体生成受损
应激颗粒是在细胞应激作用下形成的无膜结构。在NIID细胞模型中可以检测到应激颗粒的形成。在基础条件下,转染uN2C-(GGC)90-mCherry质粒的HEK293T细胞比转染uN2C-(GGC)20-mCherry质粒的HEK293T细胞以及未转染的细胞均表现出更多G3BP1阳性的应激颗粒。然而,应激颗粒的诱导剂处理后,三组间出现应激颗粒的细胞比例无明显差异。这些结果表明NOTCH2NLC GGC重复序列在基础条件下可诱导NIID细胞模型中应激颗粒的形成。同时免疫印迹结果也表明NOTCH2NLC GGC重复序列可以影响蛋白质的合成。

图6 NOTCH2NLC GGC重复序列在NIID细胞模型中可诱导应激颗粒的增强形成,抑制全细胞翻译